在阅读此文之前,辛苦您点击一下“关注”,既方便您进行讨论和分享,又能给您带来不一样的参与感,感谢您的支持!

宾夕法尼亚大学米娅·莱文实验室,最近在《Science》发了篇有意思的研究。

他们盯着果蝇的端粒保护蛋白琢磨,居然解开了生物学上一个挺矛盾的事儿那些维持生命的关键蛋白,按理说该很稳定,偏偏在进化中变得飞快。
这事儿得从基因组里的"内战"说起,细胞里可不是风平浪静的,有些叫转座子的遗传元件特自私,老想着在基因组里扩张地盘。
就像果蝇里的HeT-A、TART这些逆转录转座子,没事就往染色体末端插一脚。
宿主细胞当然不能坐视不管,端粒保护系统就是专门对付它们的防御部队。
我觉得这个"红皇后效应",用来解释这事特别形象。
就像爱丽丝梦游仙境里说的,你得拼命跑才能留在原地。
转座子不停变异想突破防御,宿主蛋白也得跟着进化升级,不然就被淘汰。
这场持久战,逼着双方都不敢停下进化的脚步。
果蝇的端粒系统跟咱们人类不一样,走了个"以毒攻毒"的路子。
人类靠端粒酶延长端粒,果蝇干脆让转座子在染色体末端复制,用这些自私元件来维持染色体长度。
听起来挺冒险,但自然界就是这么奇妙,有时候歪招反而成了生存智慧。
这里就出现个大难题:负责保护端粒的HOAP和HipHop蛋白,对果蝇来说重要得要命,没它们染色体就得融合。
可测序发现,这俩蛋白在不同果蝇物种里差异大得离谱,进化速度比其他蛋白快得多。
功能这么保守,序列为啥变得这么欢?
研究团队做了个大胆实验,用CRISPR把黑腹果蝇的HipHop基因换成近亲雅库巴果蝇的版本。
结果惨不忍睹,幼虫死亡率飙升,染色体末端全乱套了。
本来以为是整个蛋白不兼容,后来发现关键就六个氨基酸在捣乱。
把黑腹果蝇HipHop的这六个位点换成雅库巴果蝇的版本,有丝分裂时染色体融合率一下子冲到93%。

反过来把雅库巴果蝇的这六个位点改回去,居然又能正常存活。
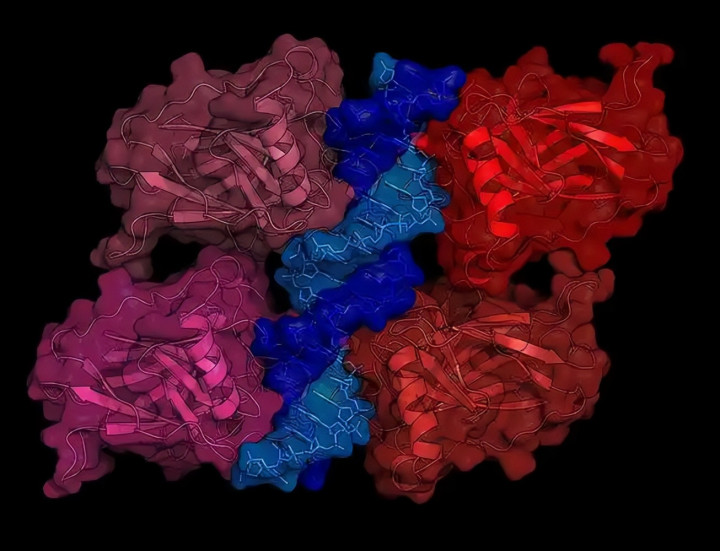
这六个氨基酸就像蛋白互作的密码,错一个都不行,解开密码的关键在结构生物学。
AlphaFold模型显示,雅库巴果蝇HipHop有个带正电的精氨酸,正好能跟自家HOAP的负电天冬氨酸形成稳定盐桥。
但黑腹果蝇HOAP没这个位点,异源HipHop来了就抓不住,等于防御系统断了电。

更绝的是"双重置换实验",同时把雅库巴果蝇的HipHop和HOAP都导进去,果蝇居然又活了。
这说明蛋白搭档就像舞伴,得一起进化才能配合默契。
一方变了,另一方必须跟着变,不然整个系统就得崩溃。
这个发现最让人意外的是杂合子的表现,按理说带一份正常基因该没事,可这些果蝇胚胎孵化率极低。
研究人员推测,异源HipHop会抢着结合父本端粒,把本该保护精子端粒的K81蛋白挤走,自己又拉不来HOAP,结果父本染色体直接裸露断裂。
果蝇胚胎发育有个"生死时速"阶段,受精后几分钟内就得完成端粒保护蛋白的交接。
这个时间窗口容错率极低,异源蛋白稍微慢点或者结合不稳,就会造成不可逆损伤。
发育早期的这种严苛筛选,其实是在加速淘汰不兼容的蛋白组合。
这正好印证了Dobzhansky-Muller模型,不同物种积累的基因突变单独看没事,碰到一起就可能致命。
HOAP和HipHop的协同进化差异,实际上成了物种间看不见的遗传屏障。
原来物种隔离不一定是染色体数目差异,几个氨基酸的变化就可能筑起高墙。
想想看,从分子错配到生殖隔离,这中间的链条居然这么清晰。
以前总觉得物种形成是个缓慢模糊的过程,现在发现可能在分子层面早就埋下了伏笔。
那些看似不起眼的蛋白互作界面,其实藏着物种分化的密码本。
这个研究彻底颠覆了,"保守功能等于保守序列"的老观念。
生命系统不是静态的精密机器,更像动态平衡的生态系统。
必需蛋白的快速进化不是bug,而是基因组应对内部冲突的自适应策略,是红皇后效应在分子层面的精彩演绎。
这让我重新思考进化的驱动力,以前总盯着外部环境压力,现在看来基因组内部的"权力斗争"可能同样重要。
转座子这些自私元件看似捣乱,实则可能是推动进化的隐形推手,逼着宿主发展出更复杂的防御机制,未来这方面研究肯定有大文章可做。
比如人类癌症里端粒异常是不是也有类似机制,能不能通过调控蛋白互作来开发新疗法?这些基础研究看似离生活很远,说不定哪天就成了医学突破的关键。
从果蝇实验室到进化论大厦,这个发现就像一块关键拼图,它告诉我们,生命的奥秘往往藏在最细微的分子互动里。
六个氨基酸的故事还在继续,而基因组暗战的更多细节,正等着我们去揭开。
